Курс лекций по планктологии (В.Б. Вербицкий)
Генезис планктона.
Вопрос о происхождении (генезисе) планктона обсуждается и работах Фореля (Forel, 1892, 1904), Везенберг-Лунда (Wesenberg-Luiid, 1910, 1921, 1926, 1930); Вольтерека (Woltereck, 1913), Везенберг-Лунда (We- senberg-Lund, 1921), Губер-Песталоцци (Huber-pestalozzi, 1938), Уэлча (Welch, 1952), Геснера (Gessner, 1955), Воронкова (1913) и др.
Первым вопрос о происхождении (генезисе) планктона поднял Форель (Forel, 1892, 1904), который под планктоном понимал совокупность организмов, взвешенных в воде пелагической области и носимых пассивно волнами или течениями, т. е. все то, что приносит сетка из тонкого шелкового сита, протянутая в воде пелагической области. Он подчеркивал замечательное однообразие пелагического сообщества пресных озер. Не только одни и те же роды, но даже одни и те же виды находят в пелагических водах всех озер континента, чтобы не сказать всего света. Это однообразие, по Форелю, вероятно, зависит от способа происхождения пелагического сообщества.
Большую роль в заселении какого-либо возникшего озера, в наличии и возобновлении пелагического населения или в появлении вновь дифференцированного вида Форель отводит заносу с помощью перелетных водоплавающих птиц. Этим способом распространения он объясняет космополитизм пелагических групп.
Происхождение пелагической фауны и флоры Форель полагает не обязательным искать в том озере, в каком они находятся, и относить это происхождение к настоящему времени или к недавно минувшим годам. Дифференциация пелагических видов, по его мнению, могла, конечно, произойти в значительном отдалении как по времени, так и по месту, в очень отдаленных озерах, может быть, в прежние геологические эпохи.
Эта дифференциация становится весьма понятной, если исходить из общего характера пелагических организмов, из их способа парить или плавать, без связи с какой-либо твердой опорой, из их космополитизма и способности их противодействовать вредным влияниям, обеспечивающей возможность переноса их или их зародышей из одного озера в другое. Всякое живое существо, обладающее всеми этими свойствами, может, если оно попадает случайно в открытое озеро, тотчас и окончательно сделаться членом пелагического сообщества.
То же самое может произойти и с любым организмом литорального сообщества, регулярно выносимым горизонтальными ветровыми течениями в пелагическую область: если они обладают способностью приспособления к новым жизненным условиям, то они становятся членами пелагического сообщества.
Везенберг-Лунд (Wesenberg-Lund, 1910, 1921, 1926, 1930) считал, что родиной пресноводного планктона служат дно озера, берег и мелкие пруды, от которых организмы планктона постепенно отходили, приспосабливаясь к жизни в толще воды. Этот процесс приспособления происходил с незапамятных времен и не прекращается и в настоящее время. Если принять такое представление, говорит Везенберг-Лунд, становится понятной чрезвычайно малая родственная связь между отдельными организмами планктона и, наоборот, большая близость очень многих планктических форм к известным донным и береговым формам. Вероятно, направление развития шло от ползающей береговой фауны и медленно плавающих животных или же от сидячих водорослей к планктону пелагической области.
Некоторые из этих линий развития удается выяснить с большой долей вероятия, о других мы пока ничего еще не знаем. Из этой теории также становится понятным, почему пресноводный планктон содержит столь далеко отстоящие друг от друга формы, такие незначительные рода и семейства, часто с одним единственным видом. Они являются крайними членами рядов развития, промежуточные формы которых, соединяющие планктические элементы с донными и береговыми формами, утеряны. В пользу всего этого говорит также и наличие у многих представителей пресноводного планктона покоящихся стадий, погружающихся на глубину и сохраняющихся на дне водоема.
Многие ясно выраженные планктические организмы, как ракообразные или синезеленые водоросли, проводят большую часть своего времени в покоящихся стадиях на дне или у берега, пелагическая же стадия очень коротка. Волны, ветер и лед подымают покоящиеся стадии и выносят их в пелагическую область, где, делясь или партеногенетически размножаясь, они образуют большой максимум, сменяющийся снова стадиями покоя и погружением на дно.
Точно так же Вольтерек (Woltereck, 1913) считал, что кладоцеры произошли от ползающих форм мелких водоемов. Основанием такого допущения служит то, что их дифференциация произошла только в связи с послеледниковым образованием озер.
Способы, использованные ими для приспособления к полностью изменившемуся образу жизни, к которому они вынуждены были перейти при превращении заросшего местообитания в глубокий водоем или при переселении во вновь возникшее озеро, были разные.
Вольтерек предполагал, что ныне живущие пелагические кладоцеры происходят от четырех разных исходных форм.

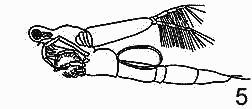
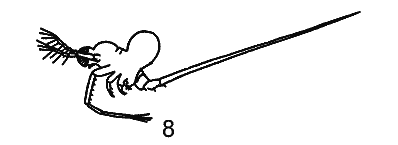
РЯД 1-й ПЕЛАГИЧЕСКИЧЕСКИХ ПРИСПОСОБЛЕНИЙ. (ПО Woltereck, 1913).
1 — Sida crystallina, в общем литоральная форма, с
прикрепительным аппаратом на спине;
2 — Diaphanosoma brachyurus всегда пелагическая, в озерах
и прудах;
3 — Holopedium gibberum, раковина изменилась в
студенистую капсулу, пелагическая форма;
4, 5 — Leptodora kindti, с удлиненным телом и
рудиментарной раковиной, пелагическая.
Первый ряд начинается от Sida crystallina - формы, обычно прикрепляющейся к листьям прибрежных растений, хотя часто свободноплавающей в пелагиали озер, и заканчивается типично пелагическими Diaphanosoma brachyurum, Ноlopedium и, вероятно, Leptodora kindti.
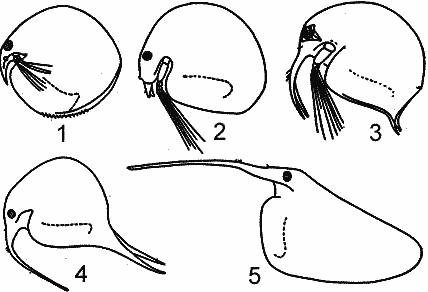
РЯД 2-й ПЕЛАГИЧЕСКИЧЕСКИХ ПРИСПОСОБЛЕНИЙ. (ПО Woltereck, 1913). |
Второй ряд начинается от прибрежно обитающего Chydoras sphaericus, который иногда появляется в пелагиали и заканчивается всегда обитающей в пелагиали Воsmina.
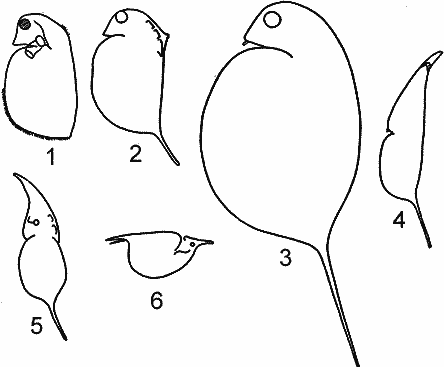
РЯД 3-й ПЕЛАГИЧЕСКИЧЕСКИХ ПРИСПОСОБЛЕНИЙ. (ПО Woltereck, 1913). |
Третий ряд имеет исходными Daphnia pulex и D. magna и заканчивается типичными пелагическими D. cucullata и Scapholeberis.
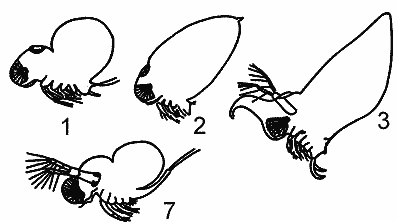 |
 |
 |
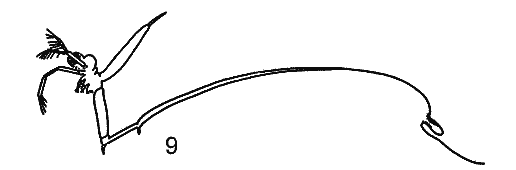 |
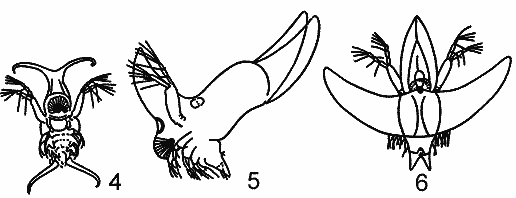
РЯД 4-й ПЕЛАГИЧЕСКИЧЕСКИХ
ПРИСПОСОБЛЕНИЙ. (ПО Woltereck, 1913).
1 — 6 — ряд podon — Evadne (с укороченным
абдоменом):
1 — p. intermedius,
2 — Evande spinifera,
3, 4 — E. hircus;
5, 6 — E. maximovitschi;
7 — 9 — ряд poluphemus — Bythotrephes (с
вытянутым абдоменом):
7 — poluphemus pediculus;
8 — Bythotrephes longimanus;
9 — Cereopagis.
Четвертый ряд состоит из полифемид, расположенных в таком порядке: podon - Evadne-Cereopagis - Bythotrephes.
Бошан (Beauchamp, 1928) и Везенберг-Лунд (Wesenberg-Lund, 1930) на примере коловраток показали, что последние первоначально были бентическими ползающими организмами, которые постепенно освоили свободную водную массу. Растительность прудов и небольших озер служила своего рода трамплином, от которого эта группа организмов оттолкнулась и достигла пелагической области.
При этом можно проследить различные направления развития от бентических до пелагических видов. Последние представляют собой конечную стадию в длинном ряду развития, ведущем от донной в пелагическую область. Коловратки, обитающие здесь, должны в большей или меньшей степени приобрести особенности, характерные для обитателей пелагиали. Эти особенности: большая прозрачность тела, приспособленная для удержания в состоянии свободного плавания без необходимости пользоваться твердой опорой.
Кроме того, для пресноводных организмов следует назвать изменчивость, резко выраженную способность привести строение своего тела и соответствие с регулярно по сезонам изменяющимися условиями существования в свободной толще воды. Эта изменчивость проявляется как локально, так и во времени.
Несомненно, это изменение формы наиболее высокого развития достигает среди пелагических, нежели среди бентических пресноводных организмов, и может считаться важным признаком планктичности подобно прозрачности и приспособлениям к уменьшению остаточного веса или к увеличению сопротивления формы.
Таким образом, коловратки подчиняются одному и тому же закону: чем больше они удаляются от бентического образа жизни к пелагическому, чем больше они становятся круглогодичными планктическими организмами, тем сильнее их изменчивость, как локальная, так и сезонная.
Что касается возраста планктона, то считается, что планктон - одно из самых старейших сообществ нашей планеты. Правда, в настоящее время трудно для этого привести удовлетворительные геологические доказательства, так как большая часть нежных оболочек из клетчатки или студенистых образований, равно как и слабо окремненных оболочек, в ископаемом состоянии не сохраняется или принадлежит к недостоверно известным.
Однако число находок из отложений более ранних геологических эпох множится и они свидетельствуют о высоком возрасте планктических организмов (в миоценовых отложениях из различных мест Европы, в меловых отложениях Саксонии, в отложениях перми и триаса Америки, юры, мела и палеогена Европы, Азии, Австралии и бывшего СССР).
Большой интерес представляет теория, поддерживаемая рядом геологов. Суть ее в том, что древние моря были значительно менее солеными, чем в настоящее время. Даже полагают, что древние внутренние воды и наиболее древние океанические воды были сходны по химическому составу и только потом постепенно с возрастом дивергировали в характере.
Если это так и если пресноводный планктон является одним из старейших сообществ на земле, тогда различные компоненты современного пресноводного планктона могли найти свой путь во внутренние воды только в период раннего возраста Земли, когда контраст между двумя категориями вод был столь незначителен, что препятствий для миграции из одной и другую не было.
Кроме того, поскольку планктон является важным звеном в пищевой цепи водных животных, то его появление в примитивных пресных водах должно было предшествовать приходу более высокоорганизованных водных животных.
Существует несколько теорий по вопросу о первоначальных путях вхождения планктона в пресные воды:
Теория, предполагающая древнее вхождение планктона в пресные воды из тропических областей - теория, которая, по-видимому, мало кем поддерживается в настоящее время.
Теория полярного происхождения пресноводного планктона, стоящая за иммиграцию арктических форм в пресные воды Европы в ледниковый период.
Теория, по которой пресноводный планктон происходит от донных и литоральных форм, приспособившихся более или менее к пелагической жизни и сделавшихся независимыми от дна и мелководья.
В связи с этой теорией утверждается, что пресноводный планктон гораздо старше ледникового возраста, что в то время как в ледниковую эпоху пресноводный планктон мог быть вытеснен из своего обширного первоначального ареала, он снова завладел старой площадью после исчезновения льда. Поэтому влияние ледниковой эпохи на историю пресноводного планктона, согласно этой теории, было лишь преходящим.
В пользу большей древности пресноводного планктона говорит наличие хорошо выраженного космополитизма, особенно среди простейших. коловраток, кладоцер и до некоторой степени среди циклопид. Эти группы животных должны считаться филогенетически древними. По мнению Брэлима (Brelim, 1930), пресноводная фауна, особенно ее космополитическая часть, более высокого филогенетического возраста, чем огромная масса морской фауны, среди которой как раз группа коловраток и кладоцер полностью или в большей части отсутствует. Так, среди кладоцер в море имеются только представители небольшой группы полифемид - podon, Evadne, а из сидид - один род penilia, среди коловраток - Synchaeta. и pseudonotholca.
Заслуживают внимания высказывания Воронкова (1913) по поводу происхождения пресноводного планктона. Воронков считал, что современный пресноводный планктон ведет свое начало от морских существ, хотя во многих случаях далеко не непосредственно.
Приспособление морских организмов к пресной воде совершалось в разные эпохи. Нельзя представить себе, что все представители пресноводного планктона возникли одновременно. Выселение из моря наблюдается и сейчас, но главным образом донных форм. Это заставляет думать, что пресноводный планктон возник от морского не непосредственно, а гораздо большую роль в этом играла донная фауна береговой полосы, представители которой уже позже - по переходе в пресные воды - изменили свой характер.
Наиболее удобные места для перехода, конечно, те, где имело место большее или меньшее опреснение моря, например: 1) устья больших тропических рек, 2) полярные моря, где благодаря таянию льда и выносу пресных вод реками происходит сильное опреснение береговых участков, 3) моря, соединенные с океаном только узким проливом и снабженные при посредстве рек массами пресной воды (Черное море). Происходит отбор наиболее эвригалинных организмов. В результате отбора в водоемах, связанных с Черным морем, встречается много выходцев, "активных переселенцев- иммигрантов". Но имеются и реликтовые формы, к каковым, например, в Ладожском и Онежском озерах относится Mysis.
О степени древности различных групп пресноводного планктона можно судить на основании степени сходства их с ближайшей родственной морской группой. Более новыми, очевидно, будут те, которые сохранили большее сходство с последней. Поэтому, например, копеподы представляются группой более молодой, нежели кладоцеры, так как пресноводные представители первых гораздо ближе к своим морским сородичам, чем пресноводные представители кладоцер к морским кладоцерам. Кроме того, наличие у копепод свободноплавающей личинки - науплиуса - и утрата таковой у кладоцер тоже говорит в пользу признания копепод за более молодую группу. Единственное исключение - присутствие науплиусовидной личинки у кладоцеры Leptodora kindti, но зато последнюю многие считают более новой, позднее остальных кладоцер приспособившейся к пресной воде.
Если отдельные формы переселялись в пресные воды разновременно, то, конечно, в прежние эпохи состав планктона мог представляться весьма отличным от современного. Прежде всего в его состав не должны были входить более новые группы, и наоборот, вероятно, входили такие, которые теперь отсутствуют. Во-вторых, древний планктон содержал большое количество свободноплавающих личинок.
Что касается того, являются ли пресноводные плапктеры прямыми потомками морских планктеров или же их происхождение может быть иным, то на этот счет существуют различные взгляды. Во всяком случае, мнение, что пресноводный планктон происходит непосредственно от морского и все озера, где имеется планктон, являются остатками морей, т. е. реликтовыми водоемами, Воронков считает неверным.
Если говорить о связи пресноводного планктона с морским, то только с неритическим планктоном, с которым наблюдается большое сходство. Так, например, две главные группы лимнопланктона - коловратки и кладоцеры - в море живут только в неритической зоне. Однако новые формы не столько создаются за счет неритического планктона, сколько являются потомками обитателей дна, например, Difflugia limnetica.
Словом, весьма вероятно, что представители современных планктических организмов, происходя в конечном счете из моря, далеко не всегда имеют своими предками морских планктеров, и наоборот, во многих случаях они возникли и не от планктеров и не как планктеры. И в пресной воде, как и в морской, береговая зона и эстуарии рек - места творчества новых форм.
